目的 探讨脂多糖(LPS)预处理对幼鼠癫痫发作及海马炎症介质Toll样受体4/高迁移率族蛋白1/磷酸化核因子抑制蛋白α(TLR4/HMGB1/P-IκB-α)表达的影响。方法 出生21 d的SD鼠随机分为生理盐水组(对照组),模型Ⅰ组和模型Ⅱ组,模型Ⅰ组采用海人酸(KA)诱导癫痫发作,模型Ⅱ组在应用KA前2 h腹腔注射LPS,观察幼鼠癫痫发作的行为学表现,荧光定量PCR检测癫痫持续状态(SE)后3 h和24 h各组海马TLR4和HMGB1基因的表达,Westernblot法检测SE后3 h、24 h各组海
腹腔注射脂多糖对幼鼠癫痫发作及海马TLR4/HMGB1/P-IκB-α表达的影响
禚志红1) 张 静1) 李国栋2) 王怀立1)△
1)郑州大学第一附属医院儿科,河南 郑州 450052 2)郑州大学生命科学学院,河南 郑州 450052
基金项目:河南省高等学校重点科研项目计划(13A320433);郑州大学第一附属医院“院内青年创新基金计划项目”
△通信作者:王怀立(1965-),男,博士,主任医师。研究方向:小儿神经及重症医学。Email:whlek6527@126.com
【摘要】 目的 探讨脂多糖(LPS)预处理对幼鼠癫痫发作及海马炎症介质Toll样受体4/高迁移率族蛋白1/磷酸化核因子抑制蛋白α(TLR4/HMGB1/P-IκB-α)表达的影响。方法 出生21 d的SD鼠随机分为生理盐水组(对照组),模型Ⅰ组和模型Ⅱ组,模型Ⅰ组采用海人酸(KA)诱导癫痫发作,模型Ⅱ组在应用KA前2 h腹腔注射LPS,观察幼鼠癫痫发作的行为学表现,荧光定量PCR检测癫痫持续状态(SE)后3 h和24 h各组海马TLR4和HMGB1基因的表达,Westernblot法检测SE后3 h、24 h各组海马TLR4、HMGB1、P-IκB-α蛋白的表达。结果 与对照组相比:模型Ⅰ组、模型Ⅱ组海马TLR4基因在SE后3 h表达均显著增加(t=4.806,P<0.05;t=4.954,P<0.05),SE后24 h也显著增加(t=3.924,P<0.05;t=3.792,P<0.05),模型Ⅰ组、模型Ⅱ组海马TLR4蛋白表达在SE后3 h均显著增加(t=7.804,P<0.05;t=8.385,P<0.05),SE后24 h也显著增加(t=4.256,P<0.05;t=4.262,P<0.05);模型Ⅰ组、模型Ⅱ组海马HMGB1基因在SE后3 h表达均显著增加(t=3.626,P<0.05;t=5.255,P<0.05),SE后24 h也显著增加(t=4.046,P<0.05;t=2.836,P<0.05),模型Ⅰ组、模型Ⅱ组海马HMGB1蛋白在SE后3 h均无显著性增加(t=0.389,P>0.05;t=0.213,P>0.05),SE后24 h也无显著性增加(t=0.106,P>0.05;t=0.279,P>0.05)。模型Ⅰ组、模型Ⅱ组海马P-IκB-α蛋白表达在SE后3 h均显著增加(t=4.383,P<0.05;t=6.627,P<0.05),SE后24 h也显著增加(t=14.521,P<0.05;t=19.458,P<0.05)。模型Ⅱ组与模型Ⅰ组相比,LPS预处理可显著增加海马TLR4基因在SE后3 h的水平(t=2.362,P<0.05),及SE后3 h TLR4蛋白表达(t=4.284,P<0.05);且显著增加SE后3 h、24 h P-IκB-α蛋白表达(t=4.249,P<0.05;t=9.120,P<0.05);但对SE后3 h、24 h HMGB1基因水平无显著性影响(t=0.569,P>0.05;t=0.691,P>0.05),对SE后3 h、24 h HMGB1蛋白表达也无显著性影响(t=0.168,P>0.05;t=0.385,P>0.05)。结论 LPS预处理加重幼鼠癫痫发作,使TLR4/P-IκB-α表达升高,对HMGB1表达无显著改变。
【关键词】 脂多糖;海人酸;癫痫;高迁移率族蛋白1;Toll样受体4;P-IκB-α;大鼠
【中图分类号】 R-332 【文献标识码】 A 【文章编号】 1673-5110(2018)02-0117-07 DOI:10.12083/SYSJ.2018.02.031
The influnces of Intraperitoneal injection of LPS on infant rats epileptic seizures and expression of TLR4/HMGB1/P-IκB-α in Hippocampus
ZHUO Zhihong1),ZHANG Jing1),LI Guodong2),WANG Huaili1)
1)Department of Pediatric,The First Affiliated Hospital of Zhengzhou university,Zhengzhou 450052,China;2)School of Life Sciences,Zhengzhou University,Zhengzhou 450052,China
【Abstract】 Objective To investigate the effect of pretreatment with lipopolysaccharide (LPS) on infant rats epileptic seizures and the expression of inflammatory mediators such as TLR4/HMGB1/P-IκB-αin hippocampus.Methods The rats at postnatal 21th day (21d) were randomly divided into saline control group (control group),model I and model II group.The model groups were induced into epilepsy model by kainic acid (KA),and the model II group were pretreated with intraperitoneal injection of LPS 2 hours before KA application.Behavior of infant rats with epilepsy seizures were observed and given scores according to Racine score,and the gene expression of TLR4 and HMGB1 at 3h and 24h after SE were detected by fluorescence quantitative PCR,and the protein expression of TLR4,HMGB1,P-IκB-α at 3h and 24h after SE were detected by western blot.Results Compared to control group,the TLR4 gene of model group Ⅰ and model group Ⅱ at 3th hour after SE in hippocampus were significantly increased (t=4.806,P<0.05;t=4.954,P<0.05),and also increased significantly at 24th hour after SE (t=3.924,P<0.05;t=3.792,P<0.05),and the TLR4 protein expression level of model group Ⅰ and model group Ⅱ at 3th hour after SE in hippocampus were significantly increased(t=7.804,P<0.05;t=8.385,P<0.05),and also increased significantly at 24th hour after SE (t=4.256,P<0.05;t=4.262,P<0.05).At the same time,the HMGB1 gene of model group Ⅰ and model group Ⅱ at 3th hour after SE in hippocampus were significantly increased(t=3.626,P<0.05;t=5.255,P<0.05),and also increased significantly at 24th hour after SE (t=4.046,P<0.05;t=2.836,P<0.05),but the HMGB1 protein expression level of model group Ⅰ and model group Ⅱ were neither significantly increased at 3th hour after SE(t=0.389,P>0.05;t=0.213,P>0.05) nor significantly increased at 24th hour after SE(t=0.106,P>0.05;t=0.279,P>0.05).What's more,the P-IκB-α protein expression level of model group Ⅰ and model group Ⅱ at 3th hour after SE in hippocampus were significantly increased(t=4.383,P<0.05;t=6.627,P<0.05),and also increased significantly at 24th hour after SE (t=14.521,P<0.05;t=19.458,P<0.05).When comparing model group Ⅱ with model group Ⅰ,LPS pretreatment could significantly increase TLR4 gene level in hippocampus at 3th hour after SE(t=2.362,P<0.05)and TLR4 protein expression level in hippocampus at 3th hour after SE(t=4.284,P<0.05),and the P-IκB-αprotein expression level in hippocampus at 3th and 24th hour after SE also significantly increased(t=4.249,P<0.05;t=9.120,P<0.05),but there were no significant change of HMGB1 gene level in hippocampus at 3th and 24th hour after SE(t=0.569,P>0.05;t=0.691,P>0.05),nor HMGB1 protein expression level in hippocampus at 3th and 24th hour after SE(t=0.168,P>0.05;t=0.385,P>0.05).Conclusion Infant rats seizures and expression of TLR4/P-IκB-αincreased by LPS pretreatment,without significant change of HMGB1 expression.
【Key words】 Lipopolysaccharide;Kainic acid;Epilepsy;High mobility group protein 1;Toll-like receptor 4;P-IκB-α;Rat
癫痫是儿童神经系统常见的慢性疾病,约30%的患儿发展为难治性癫痫,药物治疗困难。近年来研究发现,炎症在癫痫发作的发生及维持方面发挥着复杂而重要的作用,深入研究炎症与癫痫发生发展的关系,有可能为难治性癫痫的病因治疗提供一定的线索。本研究在海人酸(kainic acid,KA)诱导幼年大鼠癫痫发作前2 h腹腔注射脂多糖(lipopolysaccharide,LPS)预处理,检测急性期海马组织Toll样受体4(Toll like receptor 4,TLR4)、高迁移率族蛋白(High mobility group protein 1,HMGB1)的基因及蛋白、磷酸化核因子抑制蛋白α(phosphorylated IkappaB-alpha,P-IκB-α)的蛋白表达水平,探讨脂多糖预处理对幼鼠癫痫发作的影响以及可能的机制。
1 材料与方法
1.1 实验动物及主要试剂仪器
1.1.1 实验动物:出生21 d的清洁级健康雄性SD大鼠,体质量40~50 g,由河南省实验动物中心提供,许可证号:SCXK(豫)2015-0004。随机分为生理盐水组7只(对照组)、海人酸诱导组26只(模型Ⅰ组)、LPS预处理组31只(模型Ⅱ组)。每组造模成功后且存活的动物再随机分为3 h、24 h两个亚组,对照组仅设3 h亚组,每个亚组7只。
1.1.2 主要试剂:KA(美国santacruz,SC-200454A);LPS(美国Sigma,DG-L2880);RNA提取试剂盒(上海生工生物工程有限公司,SK1312);RNA 逆转录试剂盒(美国Thermo scientific,K1621);荧光定量PCR预混液(美国Thermo scientific,K0251);全蛋白提取试剂盒(上海生工生物工程有限公司,BSP003);改良型BCA法蛋白质浓度测定试剂盒(上海生工生物工程有限公司,SK3051);磷酸酶抑制剂复合物Ⅲ(上海生工生物工程有限公司,PL019);抗β-actin兔多克隆抗体(中国BBI,AB10024);抗TLR4兔多克隆抗体(英国abcam,ab13556);抗P-IκB-α小鼠多克隆抗体(美国Santacruze,sc-8404);抗HMGB1兔多克隆抗体(英国abcam ab191583);山羊抗小鼠IgG (中国BBI,D110087);猴抗兔IgG(中国BBI,AB10056)。
1.1.3 主要仪器:超微量分光光度计(美国Thermo scientific,NANODROP 2000);荧光定量PCR仪(瑞士Roche,LightCycler 480Ⅱ);台式高速冷冻离心机(美国Sigma,1-14K);酶标仪(美国Molecular Devices,SpectraMax i3x);垂直电泳槽(美国Biorad,Mini-Protean 3);转移电泳槽(美国Biorad);转移电泳仪(美国Biorad,Powerpac);化学发光成像系统(美国Syngene,G:BOX Chemi-XT4)。
1.2 方法
1.2.1 实验动物模型制备及行为学观察:模型组Ⅰ及对照组腹腔注射等量的生理盐水,模型组Ⅱ腹腔注射LPS(100 μg/kg)。模型组Ⅰ和模型组Ⅱ注射生理盐水/LPS后2 h腹腔注射KA(12 mg/kg)以诱导癫痫发作,对照组注射等量的生理盐水。观察注射KA/生理盐水后2 h内各组大鼠的行为学表现,根据Racine标准对KA诱导的癫痫发作进行分级:Ⅰ级:面肌抽动,节律性咀嚼动作;Ⅱ级:节律性点头或湿狗样抖动;Ⅲ级:一侧前肢阵挛;Ⅳ级:双侧前肢阵挛并站立;Ⅴ级:全身强直阵挛、跌倒[1]。Ⅳ级及以上为造模成功,SE持续2 h后,将水合氯醛(350 mg/kg)腹腔注射使发作终止,未达到Ⅳ级者从模型组中剔除。SE后3 h、24 h留取脑组织备用。
1.2.2 荧光定量PCR法检测海马内TLR4基因和HMGB1基因:①引物的设计与合成:根据Gene Bank中 SD大鼠TLR4、HMGB1及β-actin的基因序列,Primer Primer5.0软件设计引物序列(见表1),由上海生工生物工程有限公司合成。引物用DEPC水配制为100 μM的工作液,-20 ℃保存备用。②总RNA的提取和CDNA的合成:在相应时间点将大鼠处死,冰上操作分离海马组织,按说明书提取海马组织中的总RNA;用Thermo Scientific Nano Drop2000分光光度计测定RNA的浓度和纯度。依说明书进行mRNA的反转录、合成CDNA,-20 ℃保存备用。③Q-PCR反应:参照Thermo Scientific Maxima SYBR Green qPCR 预混液说明书在Light Cycler 480 实时荧光定量PCR仪进行RT-PCR反应。分析熔解曲线特异性,2-△△Ct 分析法对大鼠海马组织TLR4、HMGB1基因相对表达量进行分析,得相对表达倍数为N(N =2-△△Ct),其中△△Ct=(待测组的目的基因平均Ct值-待测组管家基因平均Ct值)-(对照组的目的基因平均Ct 值-对照组管家基因平均Ct 值)。
表1 TLR4、HMGB1基因引物序列
| 基因名称 |
|
引物序列 |
| TLR4 |
上游: |
5'-CCAGAGCCGTTGGTGTATCT-3' |
| |
下游: |
5'-CAGAGCATTGTCCTCCCACT-3' |
| HMGB1 |
上游: |
5'-TATGGCAAAGGCTGACAAGG-3' |
| |
下游: |
5'-TTTCTTCGCAACATCACCAA-3' |
| β-actin |
上游: |
5'-GCCATGTACGTAGCCATCCA-3' |
| |
下游: |
5'-GAACCGCTCATTGCCGATAG-3' |
1.2.3 Westernblot检测海马内TLR4蛋白、P-IκB-α蛋白、HMGB1蛋白的表达
1.2.3.1 海马组织蛋白提取:大鼠在相应时间点断头取脑,冰上快速分离海马组织。蛋白提取按照试剂盒操作说明进行。提取蛋白后按照试剂盒说明进行蛋白定量。
1.2.3.2 SDS-PAGE凝胶电泳:将玻璃板洗净后用双蒸水浸泡冲洗。根据目的蛋白分子量选择相应浓度的下层分离胶。依次配制分离胶和5%的浓缩胶,待浓缩胶聚合后,将待测样品与上样液充分混合,99 ℃水浴5 min。加入电泳缓冲液,蛋白样品按30 μg依次上样。开始电泳,电泳时间1.5~2 h。
1.2.3.3 转膜:预先用甲醇浸泡PVDF膜15 ~ 20 min,蒸馏水漂洗,后浸泡于转膜缓冲液中。按照:负极-海绵垫-滤纸-凝胶-PVDF膜-滤纸-海绵垫-正极的顺序放置,关闭转膜夹并插入充满缓冲液的转移电泳槽中。将电泳槽置于4 ℃冰盒中,200 mA恒流转移1 h。
1.2.3.4 封闭和杂交:5%的脱脂牛奶室温封闭1~2 h。TBST振荡洗涤3次×10 min。用一抗稀释液分别配制一抗工作液β-actin (1:5 000),HMGB1 (1:700),TLR4 (1:700),P-IκB-α(1:50);将PVDF膜放入一抗工作液中4 ℃过夜。TBST振荡洗涤3次×10 min。将PVDF膜放入用5%脱脂牛奶配制的相应二抗工作液中,室温孵育1 h。TBST振荡洗涤3次×10 min。
1.2.3.5 显影反应:选择ECL发光法,采用数字凝胶成像分析系统进行扫描,使用Image-Pro Plus 5.0软件分析条带光密度,将目的蛋白条带的光密度值与内参(β-actin)的光密度值做比值,计算各目的蛋白的相对表达量。
1.3 统计学处理 采用SPSS 17.0软件进行统计分析,计量资料以均数±标准差(x±s)表示,2组比较采用t检验,多组间比较采用One-way ANOVA法分析。P<0.05 为差异具有统计学意义。
2 结果
2.1 各组幼鼠癫痫发作成功及死亡情况比较 行为学观察模型Ⅰ组共26只SD鼠达SE并存活16只,模型Ⅱ组共31只SD鼠达SE并存活17只。模型Ⅰ组和模型Ⅱ组大鼠在KA注射后2 h内均出现行为改变,容易激惹、攻击性强、饮食减少。见表2。
表2 各组幼鼠癫痫发作成功及死亡情况比较
| 组别 |
n |
SE且存活(只) |
急性期死亡(只) |
病死率/% |
SE成功率/% |
| 对照组 |
7 |
0 |
0 |
0 |
0 |
| 模型Ⅰ组 |
26 |
16 |
8 |
30.8 |
92.3 |
| 模型Ⅱ组 |
31 |
17 |
14 |
45.2 |
100 |
2.2 荧光定量PCR检测急性期海马组织TLR4基因、HMGB1基因的表达 模型Ⅰ组、Ⅱ组在SE后3 h、24 h海马TLR4及HMGB1基因表达量显著高于对照组。SE后3 h模型Ⅱ组TLR4表达量显著高于Ⅰ组,SE后24 h无显著性差异;SE后3 h、24 h模型Ⅰ组HMGB1表达量与模型Ⅱ组比较无显著性差异。见表3、4。
表3 各组SE后3 h和24 h海马TLR4基因表达倍数比较 (x±s)
| 组别 |
n |
TLR4 Ct(次) |
β-actin Ct(次) |
2-△△CT |
| 对照组 |
7 |
25.30±0.66 |
14.40±0.54 |
1.00±0.00 |
| 模型Ⅰ组(3 h) |
7 |
24.80±0.90 |
15.17±0.24 |
2.09±0.65a1 |
| 模型Ⅱ组(3 h) |
7 |
24.86±0.55 |
15.39±0.36 |
3.34±1.42a2 b1 |
| 模型Ⅰ组(24 h) |
7 |
25.12±0.46 |
15.38±0.28 |
1.93±0.58a3 |
| 模型Ⅱ组(24 h) |
7 |
25.39±0.55 |
15.07±0.16 |
1.74±0.48a4 c1 |
| F 值 |
|
|
|
10.479 |
| P 值 |
|
|
|
<0.05 |
注:SE:癫痫持续状态,Ct:荧光阈值循环数,TLR4:Toll样受体4,对照组2-△△CT均为1,采用SPSS17.0软件进行t检验,与生理盐水比较:t=4.806,a1P<0.05;t=4.954,a2P<0.05;t=3.924,a3P<0.05;t=3.792,a4P<0.05;与模型Ⅰ组(3 h)比较:t=2.362,b1P<0.05;与模型Ⅰ组(24 h)比较:t=0.589,c1P>0.05
表4 各组SE后3 h和24 h海马HMGB1基因表达倍数比较 (x±s)
| 组别 |
n |
HMGB1 Ct(次) |
β-actin Ct(次) |
2-△△CT |
| 对照组 |
7 |
17.24±0.50 |
14.40±0.54 |
1.00±0.00 |
| 模型Ⅰ组(3 h) |
7 |
17.36±0.97 |
15.17±0.24 |
1.62±0.42a5 |
| 模型Ⅱ组(3 h) |
7 |
17.79±0.79 |
15.39±0.36 |
1.74±0.35a6 b2 |
| 模型Ⅰ组(24 h) |
7 |
17.25±0.58 |
15.38±0.28 |
1.51±0.31a7 |
| 模型Ⅱ组(24 h) |
7 |
17.27±0.66 |
15.07±0.16 |
1.38±0.33a8 c2 |
| F 值 |
|
|
|
4.89 |
| P 值 |
|
|
|
<0.05 |
注:SE:癫痫持续状态,Ct:荧光阈值循环数,HMGB1:高迁移率族蛋白1,对照组2-△△CT均为1,采用SPSS17.0软件进行t检验,与生理盐水比较:t=3.626,a5P<0.05,t=5.255,a6P<0.05,t=4.046,a7P<0.05,t=2.836,a8P<0.05;与模型Ⅰ组(3 h)比较:t=0.569,b2P>0.05;与模型Ⅰ组(24 h)比较:t=0.691,c2P>0.05
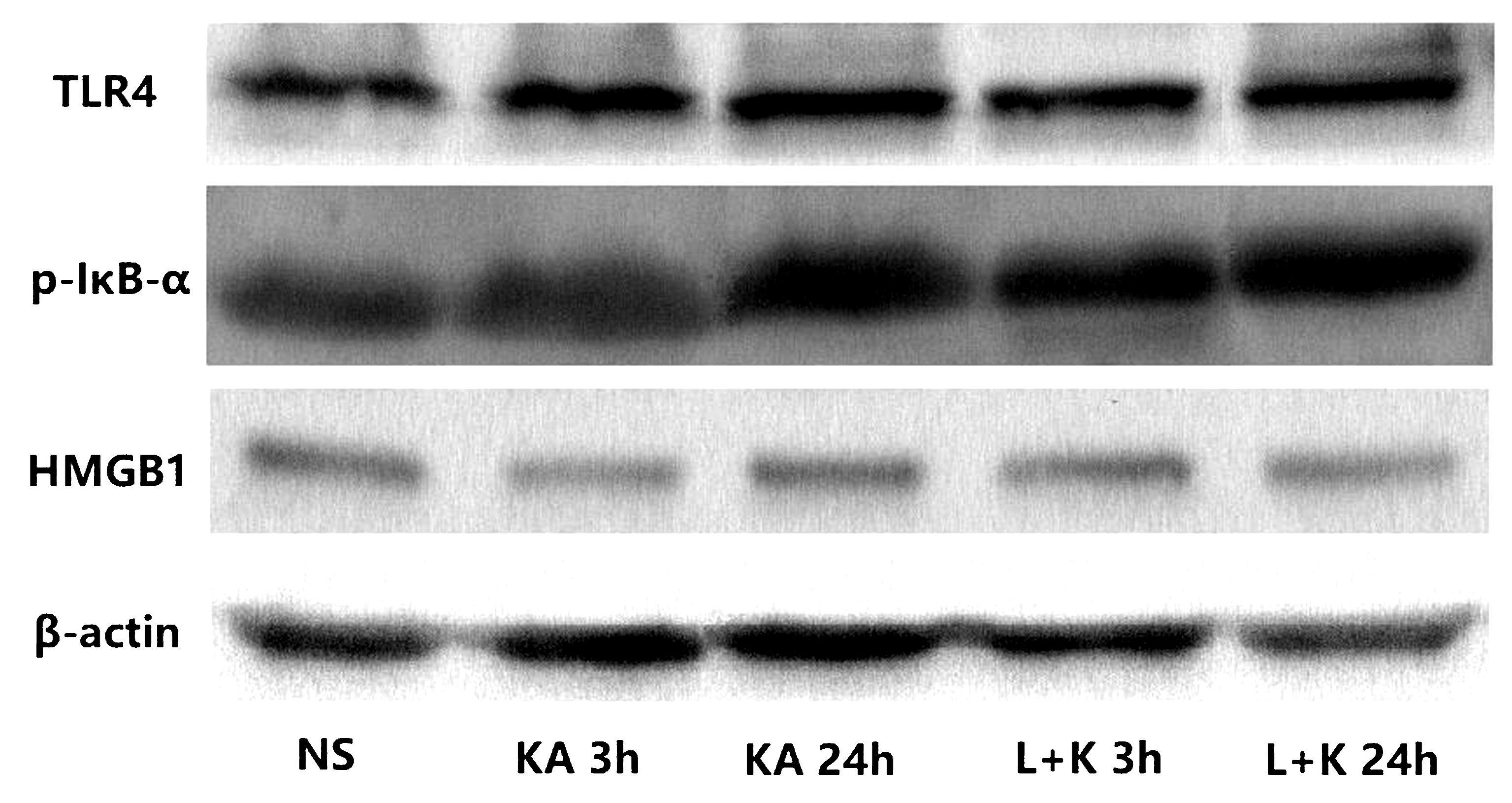
2.3 各组SE后3 h、24 hTLR4蛋白、HMGB1蛋白、P-IκB-α蛋白条带灰度值分析 Westernblot法检测各组海马TLR4蛋白、HMGB1蛋白、P-IκB-α蛋白表达 SE后各个时间点模型Ⅰ组与Ⅱ组海马TLR4蛋白与P-IκB-α蛋白表达量均显著高于对照组,与对照组比较,模型组HMGB1蛋白各时间点表达量无显著性增加;SE后3 h模型Ⅱ组TLR4蛋白表达量显著高于Ⅰ组,SE后24 h无显著性差异;SE后3 h、24 h 模型Ⅱ组P-IκB-α蛋白表达量显著高于Ⅰ组;模型Ⅰ组、Ⅱ组HMGB1蛋白表达量在各时间点均无显著性差异。采用SPSS 17.0软件进行t检验,结果见图1、表5。
图1 各组SE后3 h、24 hTLR4蛋白、HMGB1蛋白、P-IκB-α蛋白、β-actin蛋白表达情况
表5 各组SE后3 h、24 hTLR4蛋白、HMGB1蛋白、P-IκB-α蛋白条带灰度值分析 (x±s)
| 组别 |
TLR4 |
P-IκB-α |
HMGB1 |
| 对照组 |
0.16±0.075 |
0.62±0.023 |
0.333±0.95 |
| 模型Ⅰ组(3 h) |
0.61±0.032d1 |
0.87±0.021e1 |
0.41±0.93f1 |
| 模型Ⅰ组(24 h) |
0.55±0.112d2 |
1.42±0.031e2 |
0.31±0.99f2 |
| 模型Ⅱ组(3 h) |
0.84±0.086d3,g1 |
1.32±0.044e3,g3 |
0.37±0.99f3,g5 |
| 模型Ⅱ组(24 h) |
0.63±0.094d4,g2 |
2.13±0.035e4,g4 |
0.40±0.98f4,g6 |
| F值 |
11.384 |
88.344 |
0.567 |
| P值 |
<0.01 |
<0.01 |
>0.05 |
注:SE:癫痫持续状态,TLR4:Toll样受体4,P-IκB-α:磷酸化核因子抑制蛋白α,HMGB1:高迁移率族蛋白1,采用SPSS17.0软件进行t检验,与生理盐水比较:t=7.804,d1P<0.05,t=4.256,d2P<0.05,t=8.385,d3P<0.05,t=4.262,d4P<0.05;t=4.383,e1P<0.05,t=14.521,e2P<0.05,t=6.627,e3P<0.05,t=19.458,e4P<0.05;t=0.389,f1P>0.05,t=0.106,f2P>0.05,t=0.213,f3P>0.05,t=0.279,f4P>0.05。与模型Ⅰ组比较:t=4.284,g1P<0.05,t=0.737,g2P>0.05,t=4.249,g3P<0.05,t=9.120,g4P<0.05,t=0.168,g5P>0.05,t=0.385,g6P>0.05
3 讨论
近年来的实验和临床研究提示,炎症反应可能参与癫痫的发生发展,但大多数数据来源于成年癫痫患者或成年啮齿动物,对儿童癫痫患者或幼年动物的研究较少。脑内炎症反应与癫痫发作可能互为因果关系:(1)癫痫发作和继发细胞损伤可引发炎症反应;(2)炎症反应可能促进癫痫发作;(3)炎症反应可能参与癫痫发作后持久的细胞损伤[2-6]。针对炎症反应的治疗方法对一些难治性癫痫及癫痫持续状态的治疗效果也优于传统抗癫痫药[6]。LPS也称内毒素,是革兰氏阴性菌细胞壁的主要成分之一,能被TLR4识别并激活胶质细胞的固有免疫反应。TLR4是第一种被认为识别病原相关分子模式(Pathogen-associated molecular patterns,PAMPs)的受体,LPS可以被TLR4识别引发机体的免疫反应,从而抵抗病原体入侵[7-8]。张艳祥等[9]通过海马内注射LPS诱发大鼠癫痫发作,且通过LPS活化TLR4可以加重锂-匹罗卡品诱导的大鼠癫痫持续状态。HMGB1是一种核染色质成分,正常生理情况下存在于细胞核中,死亡的细胞释放的HMGB1被视为组织损伤的信号,可被TLR4识别并激活固有免疫反应。HMGB1引发的免疫反应与外源性病原体入侵时相似,但分子机制仍未明确[7,10-12]。正常生理条件下活化核转录因子κB(Nuclear factor-κ-genebinding,NF-κB)与调节蛋白κB抑制剂(IκB-α)结合,以失活状态存在于胞质中。当细胞受到TNF-α、LPS或吸入性麻醉剂等一系列外源性刺激时,TLR4/HMGB1信号通路依赖MyD88活化NF-κB,活化的NF-κB转移入核并促进炎症介质表达[13-14]。本研究在KA诱导幼年大鼠癫痫发作前2 h给予LPS预处理,检测急性期海马组织中TLR4、HMGB1基因及蛋白、P-IκB-α蛋白表达水平,探讨TLR4/HMGB1/IκB-α在LPS加重幼年大鼠癫痫发作中的变化。
在癫痫小鼠模型和人类颞叶癫痫、Rasmussen脑炎的研究中发现,TLR4/HMGB1通路在癫痫慢性病程中可以促进惊厥发作[15-18],也参与胶质瘤相关癫痫的发生机制[19-20]。临床研究也发现癫痫患儿血清中HMGB1等炎症因子水平升高[16,21-24]。HMGB1作为一种损伤相关分子模式(Damage-associated molecular patterns,DAMPs),正常情况下存在于胞核中,当组织损伤时由死亡的细胞释放到胞外。研究发现,海人酸诱导的癫痫细胞模型中,HMGB1可以通过改变表达量和细胞分布位置,参与癫痫发生的病理生理学过程[12,25],但是并未阐述具体机制及相关的信号通路。本研究发现LPS预处理可以增加KA造模的成功率及模型鼠的病死率,癫痫急性期海马TLR4、HMGB1的基因表达均显著增加,TLR4、P-IκB-α蛋白表达亦显著增加,但HMGB1总蛋白无显著改变;而LPS预处理可显著增加TLR4、P-IκB-α蛋白的表达,对HMGB1的基因水平及蛋白表达无显著性影响。提示LPS预处理使TLR4表达量升高,通过增加IκB-α磷酸化,在LPS预处理的幼鼠中促进癫痫发作急性期脑内炎症反应,进而加重癫痫发作。
TLR4/HMGB1信号通路可能促进惊厥的发生和持续,或许可作为治疗靶点改善难治性癫痫患者的治疗效果[8,11,26]。研究发现,通过抑制HMGB1/TLR4信号通路可以减轻癫痫发作对神经系统的炎症损伤[27],对难治性癫痫有一定的治疗效果,也可抑制自身免疫性脑脊髓炎相关的癫痫[26,28-29]。但是由于炎症网络的复杂性,需要注意一系列炎症因子的相互作用[30]。未来需要更多研究针对多种关键促炎因子联合治疗,以全面抑制炎症网络,可能达到更加一致的治疗结果。本研究结果提示、癫痫发作引起的损伤细胞释放HMGB1可以活化TLR4受体,通过磷酸化IκB-α活化下游炎症反应,LPS预处理可以加强这一现象从而加重癫痫发作,提示抑制TLR4活性或者抑制IκB-α磷酸化或许可以减轻癫痫发作引起的脑内炎症反应,进而为寻找针对病因治疗的新型抗癫痫药物提供理论依据。
4 参考文献
[1] RACINE R J.Modification of seizure activity by electri-cal stimulation.II.Motor seizure[J].Electroencephalogr Clin Neurophysiol,1972,32(3):281-294.
[2] 张静,禚志红,王怀立.脂多糖预处理对癫痫幼鼠Toll样受体4/高迁移率族蛋白1基因表达及海马硬化的影响[J].中华实用儿科临床杂志,2016,31(21):1 658-1 662.
[3] DUPUIS N,AUVIN S.Inflammation and epilepsy in the developing brain:clinical and experimental evidence[J].CNS Neurosci Ther,2015,21(2):141-151.
[4] LEGIDO A,KATSETOS C D.Experimental studies in epilepsy:immunologic and inflammatory mechanisms[J].Semin Pediatr Neurol,2014,21(3):197-206.
[5] VEZZANI A,VIVIANI B.Neuromodulatory properties of inflammatory cytokines and their impact on neuronal excitability[J].Neuropharmacology,2015,96(Pt A):70-82.
[6] MARCHI N,GRANATA T,JANIGRO D.Inflamma-tory pathways of seizure disorders[J].Trends Neurosci,2014,37(2):55-65.
[7] BIANCHI M E,MANFREDI A A.Dangers In and Out[J].Science,2009,323(5 922):1 683-1 684.
[8] FALIP M,SALAS-PUIG X,CARA C.Causes of CNS inflammation and potential targets for anticonvulsants[J].CNS Drugs,2013,27(8):611-623.
[9] 张艳祥,迟兆富,刘学伍,等.海马内注射脂多糖导致痫性发作及其机制[J].山东大学学报(医学版),2012,50(9):1-5.
[10] MISKIN C,HASBANI D M.Status epilepticus:immunologic and inflammatory mechanisms[J].Semin Pediatr Neurol,2014,21(3):221-225.
[11] MATIN N,TABATABAIE O,FALSAPERLA R,et al.Epilepsy and innate immune system:A possible immunogenic predisposition and related therapeutic implications[J].Hum Vaccin Immunother,2015,11(8):2 021-2 029.
[12] HUANG J S,WU Y,HUANG Q,et al.Expression level and distribution of HMGB1 in Sombati's cell model and kainic acid-induced epilepsy model[J].Eur Rev Med Pharmacol Sci,2015,19(15):2 928-2 933.
[13] LUO L,JIN Y,KIM I D,et al.Glycyrrhizin suppresses HMGB1 inductions in the hippocampus and subsequent accumulation in serum of a kainic acid-induced seizure mouse model[J].Cell Mol Neurobiol,2014,34(7):987-997.
[14] WANG D,SHI J,LV S,et al.Artesunate Attenuates Lipopolysaccharide-Stimulated Proinflammatory Responses by Suppressing TLR4,MyD88 Expression,and NF-kappaB Activation in Microglial Cells[J].Inflammation,2015,38(5):1 925-1 932.
[15] VEZZANI A.Epilepsy and inflammation in the brain:overview and pathophysiology[J].Epilepsy Curr,2014,14(1 Suppl):3-7.
[16] YANG W,LI J,SHANG Y,et al.HMGB1-TLR4 Axis Plays a Regulatory Role in the Pathogenesis of Mesial Temporal Lobe Epilepsy in Immature Rat Model and Children via the p38MAPK Signaling Pathway[J].Neurochem Res,2017,42(4):1 179-1 190.
[17] LUAN G,GAO Q,ZHA F,et al.Upregulation of HMGB1,toll-like receptor and RAGE in human Rasmussen's encephalitis[J].Epilepsy Res,2016,123:36-49.
[18] 李晓辉,王新军,杨如意,等.颞叶难治性癫痫患者致痫灶脑组织中HMGB1、TLR4表达及意义[J].实用医学杂志,2016,32(19):3 223-3 227.
[19] 周少龙,王新军,付旭东,等.伴有癫痫的人脑胶质瘤中HMGB1/TLR4的表达研究[J].中华神经医学杂志,2015,14(6):563-566.
[20] 王新军,杨如意,单峤,等.人脑胶质瘤组织HMGB1表达与胶质瘤相关性癫痫关系的探讨[J].中华肿瘤防治杂志,2015,22(15):1 180-1 183.
[21] 王怀立,郭庆敏,禚志红,等.癫(痫)患儿血清高迁移率族蛋白1及白细胞介素-6水平变化的意义[J].中华实用儿科临床杂志,2015,30(22):1 738-1 740.
[22] 桂兰,康靖,吕丽英.癫痫患儿血清HMGB1、IL-2、INF-γ 水平变化及其意义[J].山东医药,2017,57(11):60-62.
[23] 郭电渠,王新军,杨如意,等.癫痫患者血清HMGBI及其受体TLR4水平及临床意义分析[J].中华神经医学杂志,2016,15(9):936-940.
[24] WALKER L E,FRIGERIO F,RAVIZZA T,et al.Molecular isoforms of high-mobility group box 1 are mechanistic biomarkers for epilepsy[J].J Clin Invest,2017,127(6):2 118-2 132.
[25] KANEKO Y,PAPPAS C,MALAPIRA T,et al.Extracellular HMGB1 Modulates Glutamate Metabolism Associated with Kainic Acid-Induced Epilepsy-Like Hyperactivity in Primary Rat Neural Cells[J].Cell Physiol Biochem,2017,41(3):947-959.
[26] MAROSO M,BALOSSO S,RAVIZZA T,et al.Toll-like receptor 4 and high-mobility group box-1 are involved in ictogenesis and can be targeted to reduce seizures[J].Nat Med,2010,16(4):413-419.
[27] 金玉玲,单鹤,刘蕾.吡格列酮对戊四氮致痫大鼠Toll样受体、核因子-κB表达的影响[J].中国老年学,2016,36(16):3 890-3 893.
[28] RAVIZZA T,TERRONE G,SALAMONE A,et al.High Mobility Group Box 1 is a novel pathogenic factor and a mechanistic biomarker for epilepsy[J].Brain Behav Immun,2017 Oct 13.pii:S0889-1591(17)30464-6.
[29] LIU A H,WU Y T,WANG Y P.MicroRNA-129-5p inhibits the development of autoimmune encephalomye-litis-related epilepsy by targeting HMGB1 through the TLR4/NF-kB signaling pathway[J].Brain Res Bull,2017,132:139-149.
[30] DEY A,KANG X,QIU J,et al.Anti-Inflammatory Small Molecules To Treat Seizures and Epilepsy:From Bench to Bedside[J].Trends Pharmacol Sci,2016,37(6):463-484.
(收稿2017-12-14 修回2018-01-08)
本文编辑:张喜民
本文引用信息:禚志红,张静,李国栋,王怀立.腹腔注射脂多糖对幼鼠癫痫发作及海马TLR4/HMGB1/P-IκB-α表达的影响[J].中国实用神经疾病杂志,2018,21(2):117-123.